多倍化(基因组加倍)是物种形成和进化的重要驱动力之一,所有的被子植物在其进化史上都经历过一次或多次多倍化事件。物种染色体数目加倍可增加物种遗传多样性,产生新表型,对物种多样性的形成和维持有重要意义,但目前对物种多倍体的形成和建立机制仍知之甚少。近日,我校牧草与特色作物生物学教育部重点实验室熊志勇教授团队利用建立的芸苔属植物现代细胞遗传学染色体辨认体系,结合低覆盖全基因组重测序手段,以多倍体研究模式植物芸苔属植物油菜为研究对象,提出了植物多倍体形成的一种新机制,相关研究成果“Genome balance and dosage effect drive allopolyploid formation in Brassica”发表在国际顶级学术期刊PNAS。
目前二倍体产生未减数配子并通过三倍体桥进行基因组加倍被认为是多倍体形成的主要途径,为了模拟自然界中芸苔属异源三倍体形成,团队通过不同亲本间的杂交构建了不同类型的异源三倍体材料(图1)。
随后,团队成员利用细胞遗传学和生物信息学方法对芸苔属异源三倍体F2代植株进行了染色体精确辨认,证实了其自交F2代是由一群不同倍性的非整倍体组成,包括有近三倍体、近五倍体和近六倍体;研究发现F2代群体的染色体数目分布偏离了理论分布,染色体数目呈增加的趋势,说明F2代群体受到了一定的选择作用(图2)。
同时,团队成员还对异源三倍体F2代群体的表型、花粉活性和结实数等性状进行了分析,发现异源三倍体F2代中具有成套C基因组染色体(Complete)和剂量补偿(Compensated)的非整倍体植株具有更高的花粉活性和结实数;此外,A基因组完整(Complete)的植株,其花粉活性和结实数在整体上显著高于基因组不平衡(Imbalanced)的植株。这些结果说明基因组完整性和剂量效应对异源三倍体后代的选择导致只有基因组平衡破坏相对较小的植株才能存活(图3)。
随后,利用染色体精确识别技术连续观察异源三倍体AAC自交F2-F4代的核型变化,并结合花粉活性和结实数等性状分析,证实了芸苔属异源三倍体经过3代的自交,通过对植株的基因组平衡的选择,不断增加后代群体的染色体数目和育性,逐渐形成了新的异源四倍体;因此证明了异源三倍体可以作为异源四倍体形成的桥梁(图4)。
最后,团队成员利用生物信息学方法,对新形成的异源四倍体进行序列分析,发现新形成的异源四倍体携带有二倍体亲本A基因组染色体或染色体片段,证实了芸苔属异源三倍体介导了二倍体到异源四倍体单方向的基因流动。
该项研究证明,在基因组平衡和剂量效应的驱动下,异源三倍体以非整倍体为媒介,通过染色体数目和育性不断地增加逐步形成异源四倍体。因此,该研究发现了异源多倍体形成的新途径,提出一新的机制,并证实通过异源三倍体到异源四倍体的反复轮回不仅维持了新生异源四倍体的种间隔离还促进了二倍体向异源四倍体基因的流动。为解释新生多倍体物种的形成和维持机制提供了理论依据。
论文链接:www.pnas.org/doi/10.1073/pnas.2217672120
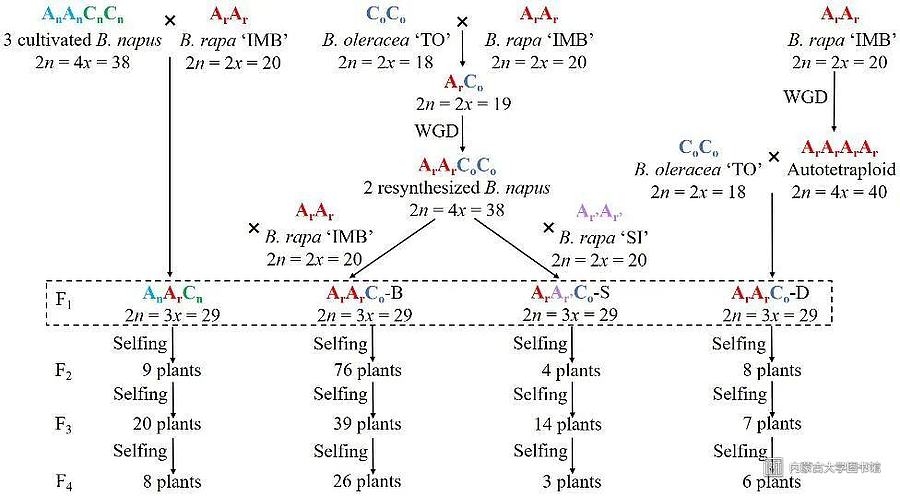
图1芸苔属异源三倍体 AAC 及其自交后代材料创建流程图
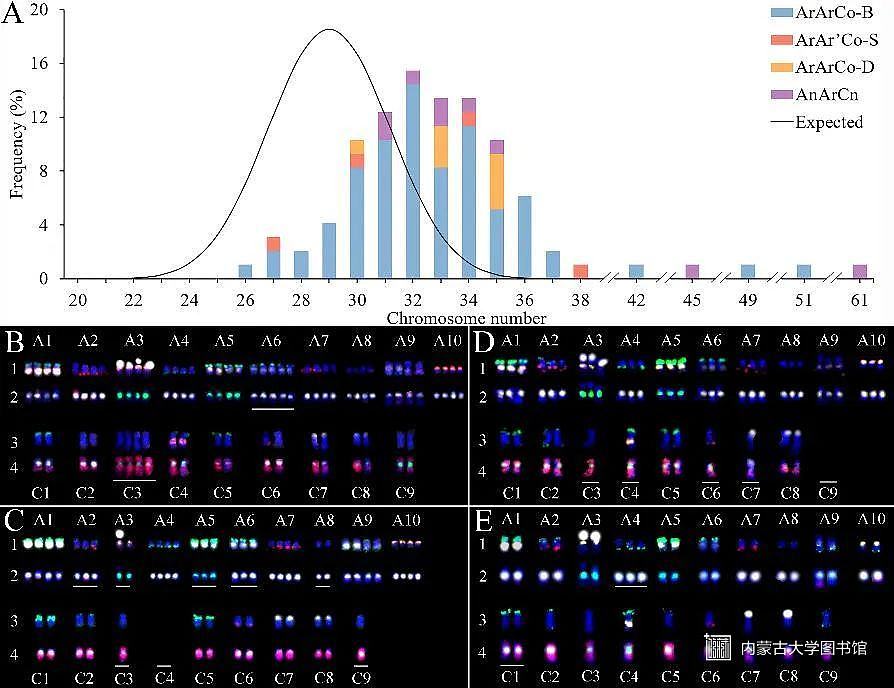
图2芸苔属异源三倍体F2代的染色体数目分布和核型分析
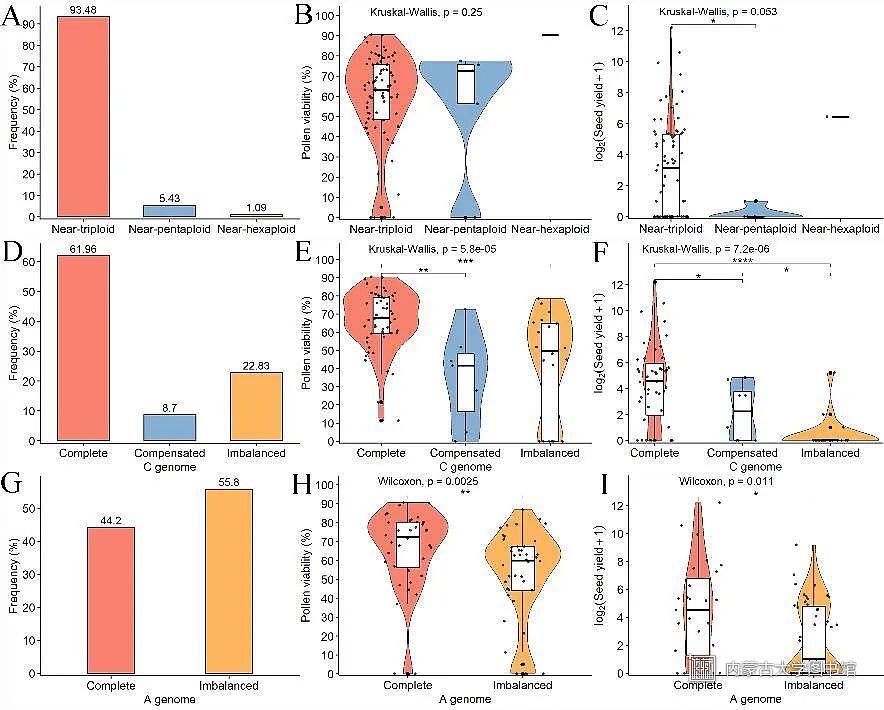
图3芸苔属异源三倍体F2代育性分析
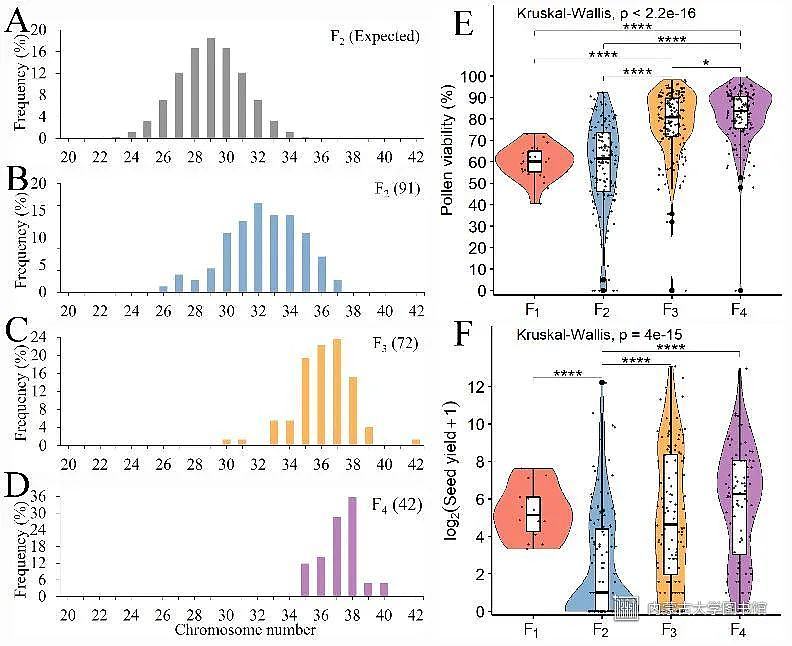
图4芸苔属异源三倍体F2-F4代群体的染色体数目分布和育性比较
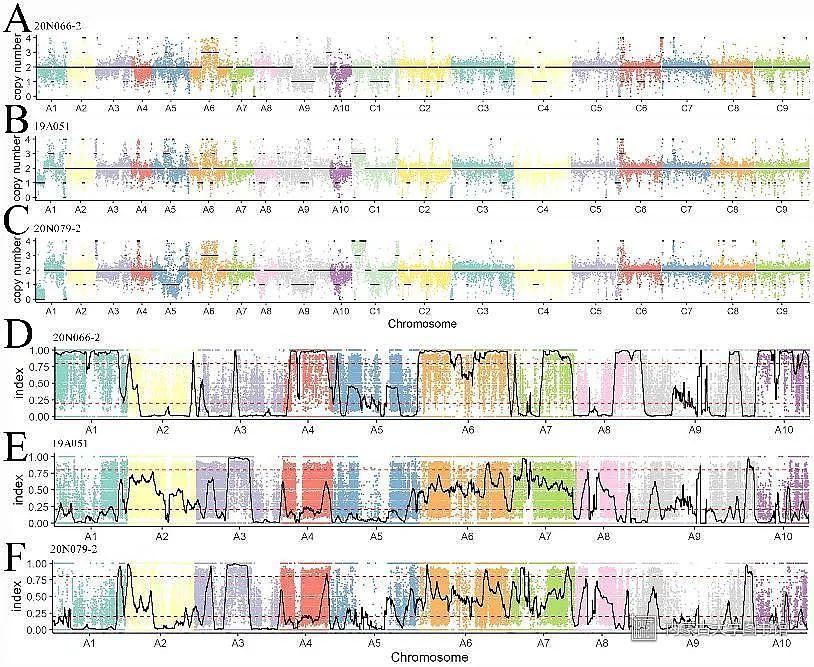
图5新形成的异源四倍体的分子核型和基因渗入分析
图1芸苔属异源三倍体 AAC 及其自交后代材料创建流程图
图2芸苔属异源三倍体F2代的染色体数目分布和核型分析
图3芸苔属异源三倍体F2代育性分析
图4芸苔属异源三倍体F2-F4代群体的染色体数目分布和育性比较
图5新形成的异源四倍体的分子核型和基因渗入分析









